VIRUS REPLICATION
Animals were first used for experimental or diagnostic work, followed by chick embryos and finally cell cultures. Numerous types of animal cell culture have found application in virology. The choice of species, tissue of origin, and type of culture (primary, cell strain, or cell line) depends on the virus and experimental objectives. Each animal virus can replicate only in a certain range of cells. Among non-susceptible cells, some have a block at an early step. (e.g. they lack suitable receptors or a factor required for expression of viral genes), so that the expression of all viral functions is prevented (resistant cells) Other cells lack a factor required for a later step, so that some, but not all viral activities are expressed (non-permissive cells). In either case, a heterokaryon formed by fusing a susceptible and a non-susceptible cell has the required functions and is usually susceptible.
Role of nucleic acid - transfection
The cells of higher organisms can be infected by naked viral nucleic acid, yielding normal virions. There are several important differences between infections by nucleic acid (transfection) and by virions.
1. The efficiency of infection with nucleic acid is much lower, by a factor of 10-6 to 10-8 in ordinary media, showing the importance of the viral coat in infectivity. The efficiency can be increased by the precipitation of viral DNA onto cells with calcium phosphate, injection or packaging into liposomes.
2. The host range is much wider with nucleic acids, which can infect resistant cells e.g. chicken cells, although resistant to poliovirus because they lack receptors for the virions, are susceptible to its RNA, but only 1 cycle of viral multiplication takes place because the progeny are again virions and cannot spread to other cells because of the lack of suitable receptors or essential factors.
3. Infectious nucleic acid can be extracted even from heat- inactivated viruses in which the protein of the capsid has been denatured; the nucleic acid can withstand much higher temperatures than the protein. The ability of nucleic acid infectivity to survive damage to the viral coat must be considered in the preparation of viral vaccines.
4. With some RNA viruses, a DNA copy of the viral RNA is infectious. e.g. polioviruses, this permits the preparation of viral genomes, such as those of vaccine strains in high quantities by avoiding the high mutation rate in replication of RNA and its lability.
5. The infectivity of nucleic acid is unaffected by virus-specific ABs, which suggests that this form of a virus could be an effective infectious agent even in the presence of immunity. However, nucleases in the body probably greatly limit its role. Of the animal viruses, papovaviruses, adenoviruses, some herpesviruses, togaviruses and picornaviruses yield infectious nucleic acid. With retroviruses, infectious DNA can be extracted from infected cells or can be made by copying the viral RNA in vitro.
Steps in Viral Replication
The following steps take place during viral replication;-
- Adsorption
- Penetration
- Uncoating
- Viral genome replication
- Maturation
- Release
1. Adsorption
The virus becomes attached to the cells, and at this stage, it can be recovered in the infectious form without cell lysis by procedures that either destroy the receptors or weaken their bonds to the virions. Animal viruses have specialized attachment sites distributed over the surface of the virion e.g. orthomyxoviruses and paramyxoviruses attach through glycoprotein spikes, and adenoviruses attach through the penton fibers. Adsorption occurs to specific cellular receptors. Some receptors are glycoproteins, others are phospholipids or glycolipids. These are usually macromolecules with specific physiological functions, such as complement receptors for EBV. Whether or not receptors for a certain virus are present on a cell depends on the species, the tissue and its physiological state. Cells lacking specific receptors are resistant. Attachment is blocked by antibodies that bind to the viral or cellular sites involved.
2. Penetration
Penetration rapidly follows adsorption, and the virus can no longer be recovered from the intact cell. The most common mechanism is receptor mediated endocytosis, the process by which many hormones and toxins enter cells. The virion is endocytosed and contained within a cytoplasmic vacuole.
3. Uncoating
A key step in uncoating is the acidification of the content of the endosome to a pH of about 5, owing to the activity of a proton pump present in the membrane. The low pH causes rearrangement of coat components, which then expose normally hidden hydrophobic sites. They bind to the lipid bilayer of the membrane, causing the extrusion of the viral core into the cytosol. For influenza virus, the acid-sensitive component is the core HA2 unit of the haemagglutinin, for adenoviruses, it is the penton base.
4. Viral Nucleic Acid Replication
Virulent viruses, either DNA and RNA, shut off cellular protein synthesis and disaggregate cellular polyribosomes, favouring a shift to viral synthesis. The mechanism of protein synthesis shut-off varies even within the same viral family. Poliovirus, using a viral protease, causes cleavage of a 200 Kd cap-binding protein, which is required for initiation of translation of capped cellular messengers. In contrast to virulent viruses, moderate viruses e.g. polyomaviruses may stimulate the synthesis of host DNA, mRNA, and protein. This phenomenon is of considerable interest for viral carcinogenesis.
DNA Viruses
With animal DNA viruses, transcription and translation are not coupled. Except for poxviruses, transcription occurs in the nucleus and translation in the cytoplasm. Generally, the primary transcripts, generated by RNA polymerase II, are larger than the mRNAs found on ribosomes, and in some cases, as much as 30% of the transcribed RNA remains untranslated in the nucleus. The viral messengers, however, like those of animal cells, are monocistronic. Transcription has a temporal organization, with most DNA viruses only a small fraction of the genome is transcribed into early messengers. The synthesis of early proteins is the key initial step in viral DNA replication. After DNA synthesis, the remainder of the genome is transcribed into late messengers. The complex viruses have immediate early genes, which are expressed in the presence of inhibitors of protein synthesis, and delayed early genes, which require protein synthesis for expression. Regulation is carried out by proteins present in the virions, or specified by viral or cellular genes, interacting with regulatory sequences at the 5' end of the genes. These sequences may respond in trans to products produced by other genes and act in cis on the associated genes. Different classes of genes may be transcribed from different DNA strands and therefore in opposite directions e.g. polyomaviruses. The transcripts may undergo post- transcriptional processing so that nonessential intervening sequences are removed.
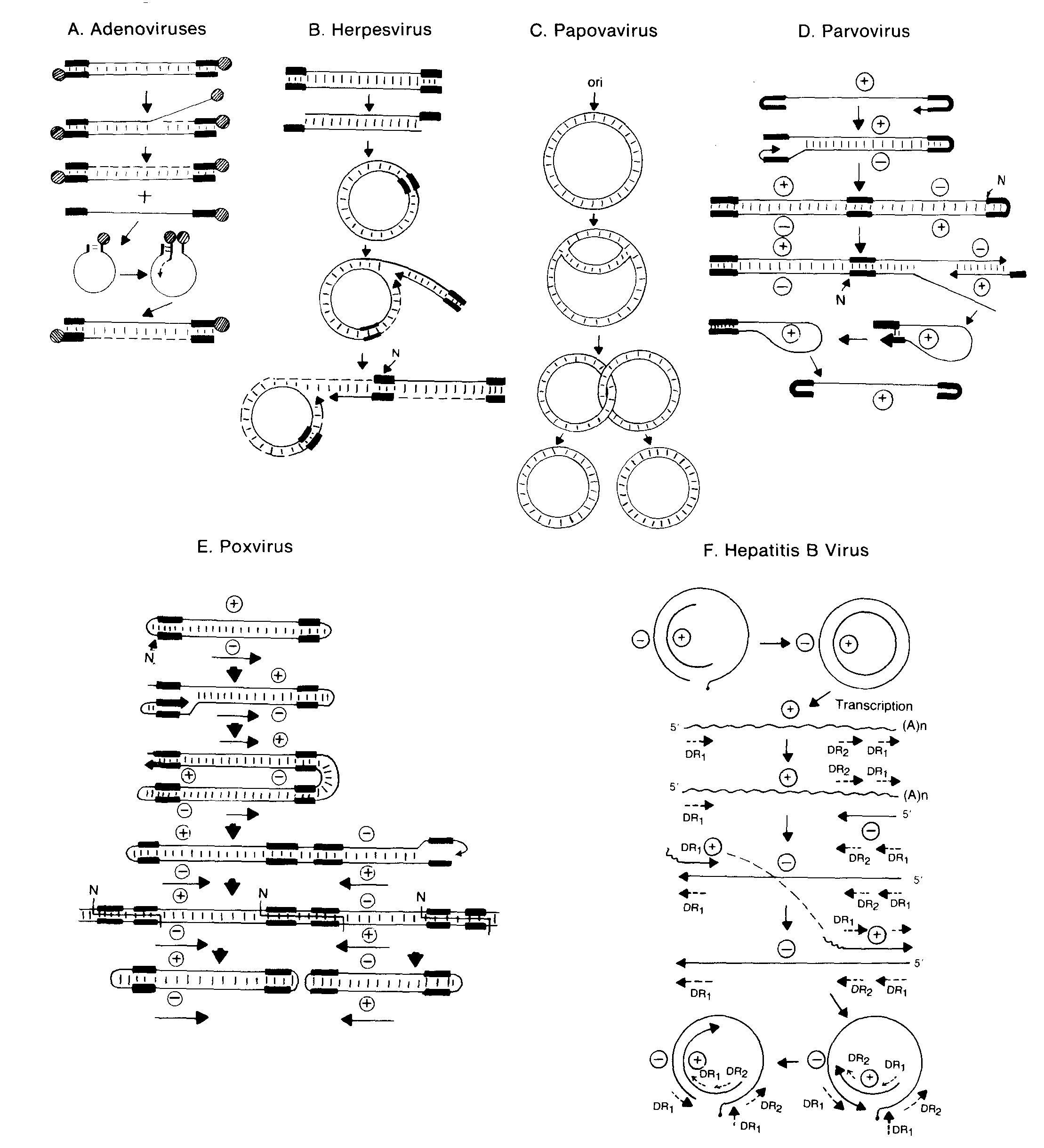
DNA replication
The mode of replication is semiconservative but the nature of the replicative intermediates depends on the manner of replication. Several methods of replication can be recognized.
A. Adenoviruses - Adenoviruses show asymmetric replication, which initiates at the 3' end of one of the strands using a protein primer. The growing strand displaces the preexisting strand of the same polarity and builds a complete duplex molecule. The displaced strand in turn replicates in a similar manner after generating a panhandle structure by pairing the inverted terminal repetitions.
B. Herpesviruses - Herpesviruses have linear genomes with terminal repeats. On reaching the nucleus, the terminal ends undergo limited exonucleotic digestion and then pair to form circles. Replication is thought to take place via a rolling circle mechanism, where concatemers are formed. During maturation, unit-length molecules are cut from the concatemers.
C. Papovaviruses - The DNA of papovaviruses are circular and the replication is bidirectional and symmetrical, via cyclic intermediates.
D. Parvoviruses - The replication of single stranded parvoviruses is initiated when +ve and -ve stranded DNA from different parvovirus particles come together to form a double stranded DNA molecule from which transcription and replication takes place.
E. Poxviruses - The striking feature of poxvirus DNA is that the two complementary strands are joined. The replicative intermediates, present in the cytoplasm, are special concatemers containing pairs of genomes connected either head to head or tail to tail.
F. Hepadnaviruses - Hepatitis B virus employs reverse transcription for replication. The genome consists of a partially double-stranded circular DNA with a complete negative strand and an incomplete positive strand. Upon entering the cell, the positive strand is completed and transcribed. RNA transcripts are in turn reverse-transcribed into DNA by a viral enzyme in several steps, following closely the model of retroviruses, including a jump of the nascent positive strand from one direct repeat (DR) to another.
RNA Viruses
The replication of RNA viral genomes is dictated by the absence of multiple translation units within the same messenger, a characteristic of all animal cell messengers. To overcome this difficulty, 3 main strategies have developed.
The viral mRNA acts directly as the messenger and is translated monocistronically, followed by cleavage to form different proteins.
The virion RNA is transcribed to yield various monocistronic mRNAs by initiating transcription at various places.
The genome itself is a collection of separate RNA fragments that are transcribed into monocistronic mRNAs.
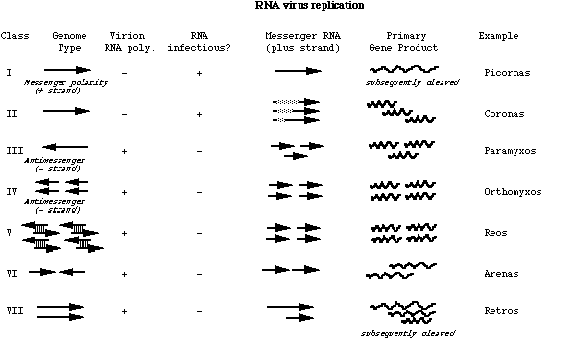
RNA viruses can be placed into 7 classes, according to the nature of the viral RNA and its relation to the messenger.
Class I (e.g. picornaviruses, flaviviruses.) The genome, having +ve polarity, itself act as the messenger, specifying information for the synthesis of both structural and nonstructural proteins. The same RNA molecule also initiate replication that requires the expression of proteins first. This format allows little control over replication e.g. Poliovirus has no independent mechanism of controlling the numbers of structural proteins made.
Class II (e.g. coronaviruses, togaviruses.) Many +stranded RNA viruses have subgenomic RNA as part of their cycle. This would allow a certain amount of control. The subgenomic mRNA cannot be recognized by the RNA polymerase. It can be used solely for the synthesis of structural proteins etc. A second way to get round the problem is to make a nested set of RNAs. The nested set of RNA is the most efficient form of control. They can control which part of their genome to express.
subgenomic PTC Nested Splicing
Picornaviridae N Y N N
Togaviridae Y Y N N
Coronaviridae Y N Y N
With togaviruses, the 49S genome RNA is first translated into polyprotein that is processed into the nonstructural proteins. The subgenomic 26S mRNA, which is transcribed from the full length -ve RNA, is translated into a smaller polyprotein that is processed into viral structural proteins. With coronaviruses, a nested set of mRNAs is generated in the following manner: the -ve transcript is first generated form the genome, which is then transcribed into monocistronic mRNAs of different sizes. Each begins with an identical short 5' leader sequence that is joined to the transcripts at the start of the various genes and continues to the 3' end of the genome. These mRNAs are not produced by splicing a genomic-size transcript because the virus is able to replicate in enucleated cells.
Class III (e.g. paramyxoviruses, rhabdoviruses.) The genome is of -ve polarity to the messenger. A virion RNA-dependent RNA transcriptase first transcribes the genomes into separate monocistronic messengers initiating at a single promoter. The transcriptase stops and restarts at each juncture between different genes.
Class IV (e.g. orthomyxoviruses, most bunyaviruses.) The -ve genome is in several distinct nonoverlapping pieces of ssRNA. The virion transcriptase generate a messenger from each piece. With orthomyxoviruses, most genomic segments contain a single gene but 2 fragments contain 2 overlapping genes: one is expressed by a full-length messenger, the other by a shorter messenger obtained from the former by splicing. The replication of orthomyxoviruses is unusual amongst RNA viruses in that it takes place within the nucleus. The nuclear function it requires is the 5' cap of cellular messengers, which it "pinches" after endonucleotic cleavage of the host messengers. The 5' cap is then used as primers in the synthesis of viral messengers.
Class V (e.g. arenaviruses, phleboviruses.) Arenaviruses have an ambisense genome in that half the genome is of -ve polarity and is transcribed into a messenger by a virion transcriptase, but the other half, which is of +ve polarity is transcribed twice: first a complete transcript of the genome is made, then the mRNA is transcribed form this transcript. This strategy is seen in the S (small) segment of the genome of phleboviruses. Ambisense genomes are unusual for RNA viruses but not for dsDNA viruses.
Class VI (e.g. reoviruses. Reoviruses.) contain distinct nonoverlapping segments of dsRNA, each is transcribed into an independent mRNA by the virion transcriptase. Most messengers are monocistronic, but one is bicistronic and expresses a second protein by initiating at an internal AUG in a different reading frame. Each segment of reovirus RNA is replicated independently. A nascent mRNA strand is first generated by the virion transcriptase, which then serves as the template for the replicase to make the negative strand. The two strands remain associated in a dsRNA molecule that ends up in a virion. This replication is asymmetric and conservative because (1) the -ve strand of the virion RNA servers as the initial template and (2) the parental RNA does not end up in the progeny.
Class VII (e.g. retroviruses.) Retroviruses are unique in that their genomes are transcribed into DNA and not RNA. They contain two identical ssRNA of +ve polarity, with a poly A tail at the 3' end and a cap at the 5' end. Each is transcribed into DNA by reverse transcriptase that then integrates into the cellular DNA as provirus. Transcription of the provirus by the cellular transcriptase yields the viral molecules that end up in virions.
Since RNA viruses of classes III to VII require a virion
transcriptase for synthesizing a messenger, their purified viral
RNAs are not infectious. Only those of classes I and II are
infectious. With RNA viruses, there is no differentiation between
early and late messengers.
Replication of Single-Stranded RNA Viruses (Classes I to V)
In all cases, replication consists of building a template strand complementary to the viral strand of the same length, which then servers as the template for progeny viral strands. These steps are carried out by a collection of enzymes of both viral and cellular origin, in association with the nucleocapsids of the infecting virions. In many instances, replication and transcription interfere with each other: with -ve stranded viruses, both template and transcripts are made from viral strands: with +ve viruses, a viral strand can be used as a messenger or replication template. Initially in the infection, there is no interference as the messenger function is needed to provide proteins needed for replication. Later the supply of these proteins regulate the rate of replication. e.g. with poliovirus, replication is initiated when the pVg protein becomes covalently linked to at the 5' ends of the RNA, apparently initiating the formation of a replication complex. Messenger and progeny often differed structurally. e.g., the messengers of influenza virus have capped leader sequences derived from cellular messengers. In addition, they lack 17 to 22 nucleotides at the 3' end. Moreover, replication requires ongoing protein synthesis to provide the required proteins, whereas transcription does not.
In RNA replication, the newly made template strand remains associated with the viral strand on which it is made, forming a double-stranded structure the length of the viral genome, known as the replicative form (RF). Synthesis of new strands occurs by conservative asymmetric synthesis, similar to adenoviruses. An RF with a nascent viral strand is known as RI (replicative intermediate) RF molecules are fairly abundant during replication because after the completion of a new strand, the replicase appear to remain associated for some time with the template before reinitiating synthesis. RFs accumulate at the end of replication, when no more RIs are formed. With the exception of orthomyxoviruses, the viral RNAs replicate in the cytoplasm. The replicase present in infected cells synthesize new viral RNA strands of both polarities. Transcription occurs at the same site as replication. It is unclear whether replication and transcription are carried out by different enzymes or by the same enzyme.
Maturation and Release
Maturation proceeds differently for naked, enveloped, and complex viruses.
Naked icosahedral viruses - Preassembled capsomers are joined to form empty capsids (procapsid) which are the precursors of virions. The assembly of capsomers to form the procapsid is often accompanied by extensive reorganization, which is revealed by changes in serological specificity and isoelectric point. eg. picornaviruses and adenoviruses. Naked icosahedral viruses are released from infected cells in different ways. Poliovirus is rapidly released, with death and lysis of infected cells. in contrast, the virions of DNA viruses that tend to mature in the nucleus tend to accumulate within infected cells over a long period and are released when the cell undergoes autolysis, and in some cases, may be extruded without lysis.
Enveloped Viruses - Viral proteins are first associated with the nucleic acid to form the nucleocapsid, which is then surrounded by an envelope. In nucleocapsid formation, the proteins are all synthesized on cytoplasmic polysomes and are rapidly assembled into capsid components. In envelope assembly, virus-specified envelope proteins go directly to the appropriate cell membrane (the plasma membrane, the ER, the Golgi apparatus), displacing host proteins. In contrast, the carbohydrates and the lipids are produced by the host cell. The viral envelope has the lipid constitution of the membrane where its assembly takes place. (eg. the plasma membrane for orthomyxoviruses and paramyxoviruses, the nuclear membrane for herpesviruses) A given virus will differ in its lipids and carbohydrates when grown in different cells, with consequent differences in physical, biological, and antigenic properties.
The envelope glycoproteins are synthesized in the following manner: the polypeptide backbone is first formed on polysomes bound to the ER, which then moves via transport vesicles to the Golgi apparatus where it attains it full glycosylation and fatty acid acylation. The matrix proteins that are present in viral envelope are usually not glycosylated and stick to the cytoplasmic side of the plasma membrane through hydrophobic domains. Matrix proteins connect the cytoplasmic domains of the envelope glycoproteins with the cell's cytoskeleton, and they gather the viral glycoproteins to form the virions. The selection of viral glycoproteins is efficient but not exclusive. eg. rhabdovirus virions contain 10 to 15% of nonviral glycoproteins. They may also contain glycoproteins specified by another virus infecting the same cell. Envelopes are formed around the nucleocapsids by budding of cellular membranes.
With orthomyxoviruses and paramyxoviruses, the viral glycoproteins incorporated in the membranes confer on the cell some properties of a giant virion. Thus cells infected by these viruses may bind RBCs (haemadsoption), and paramyxovirus-infected cells may fuse with uninfected cells to form multinucleated syncytia by the fusion of their membranes. This fusion is equivalent to the fusion of the virion's envelope with the plasma membrane of the host cell at the onset of infection.
Complex Viruses
Maturation of the highly organized poxviruses takes place in cytoplasmic foci called "factories" In contrast to simpler viruses, the poxvirus membrane contain newly synthesized lipids that differ in composition to the cellular lipids. The maturation of poxviruses after the precursors have been enclosed within the primitive membranes suggest that poxviruses may be transitional forms towards a cellular organization.
Defective Interfering Particles
Interference may occur during replication by the generation of defective interfering (DI) particles. They are formed during infection with various kinds of RNA viruses, such as rhabdoviruses, togaviruses, orthomyxoviruses, paramyxoviruses, coronaviruses and some DNA viruses (herpesviruses). With some viruses eg. VSV, the DI particles are smaller than regular particles and can therefore be obtained in pure form. They usually contain the normal virion proteins but have a shorter genome. They are replication defective and require the helper functions of a normal virus co-infecting the same cell. In early serial passages, DI particles rapidly increase in titre, then the yield of the infectious virus, and finally the total particle yield is progressively reduced.
The genomes of DI particles are internally deleted but retain both ends, which are essential for the replication of RNA viruses. With DNA viruses, the origin of replication is always conserved and often repeated. Those features show that to cause interference, the DI genomes must replicate. They deprive the regular virus of its replicase by binding to it more effectively. They do not make a replicase of their own because they are always defective in their replicase gene. The formation of DI genomes of RNA viruses is the consequence of high variability of these genomes. The DI genomes are formed by a copy choice mechanism when the replicase, having replicated part of the template, skips to another part of the same or another template. With VSV and other negatively stranded RNA viruses, 4 types of defective genomes are seen;
Deletions - the polymerase jumps to a site beyond on the same template, skipping a fragment.
Snapbacks - this occurs when the replicase, having transcribed part of the + strand, switches to the just-made - strand as template. The resultant RNA contain half+ and half- and can produce a hairpin on annealing.
Panhandle - this is formed by a similar mechanism, when the polymerase carrying a partially made - strand switches back to transcribing the extreme 5' of it, so that on annealing, the strand forms a panhandle.
Compounds - these genomes are made by a combination of deletions and snapbacks.
The competition of DI genomes with competent genomes depends not only on the structure of the DI genome but on that of the normal competent genome. Different DI genomes may interfere to very different degrees with the same competent genome and competent genomes may acquire mutations that make them resistant to the existing DI genomes. Subsequently, this is overcome by the new types of DI genomes. During viral multiplication, many types of DI genomes are continuously made and they are very heterogeneous.
Viroids
Viroids are responsible for causing serious diseases in many plants. They consist of naked RNA which does not code for any protein, nor is protein associated with it. Essentially, each viroid particle is a circular ssRNA molecule containing 250 to 400 nucleotides. They are highly resistant to enzymatic degradation because they have no free ends and because they have a very tight secondary structure (owing to self-complementary sequences). All viroid strains have similar characteristics. Their genome can be considered a dsRNA, with many unpaired short "bubbles" regions. There is no AUG initiation codon for protein synthesis, or of their complements (in case the RNA is of negative-stranded type) There is no evidence that the RNA is translated. They are replicated in the nucleus of infected cells by host enzymes through double stranded intermediates. Replication is blocked by alpha-amantine, which inhibits RNA polymerase II (the RNA polymerase responsible for generating the transcripts for mRNAs)
The base sequences of viroids have repeats, both direct and inverted, which suggest a relatedness to transposing elements. Moreover, they possess a sequence similar to that used by retroviruses. However, viroids are not transcribed into DNA, and no sequences homologous to viroids are found in the DNA of infected cells. cDNA of the viroid is also infectious and can be transcribed into regular infectious viroid particles. A striking feature of viroid RNA is the presence of sequences highly homologous to some of the small nuclear RNAs U1 and U3, which are involved in the splicing of introns in animal cells. This suggests that viroids may have originated from introns and their pathogenecity might be due to interference with the normal splicing of introns in cells. Virusoids are satellites of certain plant viruses that are encapsidated with their helper RNAs in the virions. A candidate for a viroid-like agent in humans is the delta agent which is much larger (1678bp) and is surrounded by a coat.
![]()
![]()